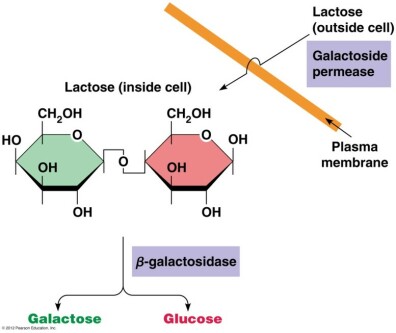
Les glucides sont la principale source d'énergie des bactéries. Le glucose est la source préférentielle mais le lactose peut également être utilisé après hydrolyse en galactose et glucose par la β-galactosidase.
L'opéron lactose est un opéron catabolique (synthèse des molécules permettant l'utilisation du lactose) qui permet de réguler l'expression de gènes impliqués dans le métabolisme du lactose.
Une petite vidéo pour se remettre en mémoire l'opéron lactose.
Rappel : Organisation de l'opéron lactose⚓
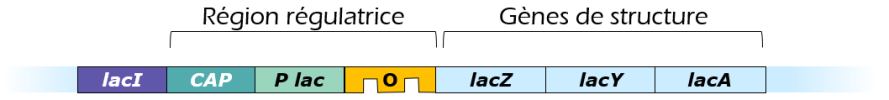
Dans l'opéron lactose, on trouve les trois gènes de structure nécessaires à l'utilisation du lactose. Ils codent pour :
La β-galactosidase (gène lacZ) qui hydrolyse la liaison β1-4 osidique des β-galactosides (donc du lactose).
La lactose perméase (gène lacY), une protéine membranaire qui permet l'entrée du lactose dans la cellule.
La thiogalactoside transacétylase (gène lacA) dont le rôle est mal connu.
Ces trois gènes de structure sont précédés par une région régulatrice qui comprend le promoteur et l'opérateur ainsi qu'un site de fixation pour la protéine CAP (= Catabolite Activator Protein, également appelée CRP pour cAMP Receptor Protein)
On trouve également en amont de l'opéron lactose, le gène régulateur (lacI) qui code pour un répresseur. Celui-ci agit en inhibant l'expression des gènes de l'opéron lactose en se liant spécifiquement sur l'ADN au niveau de l'opérateur. L'expression de ce répresseur est constitutive.
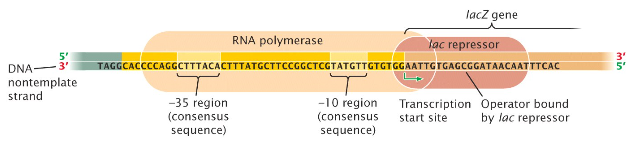
La fixation du répresseur sur l'opérateur recouvre en partie le promoteur et le gène lacZ.
Une vidéo de démonstration (donc partielle) qui montre le fonctionnement de l'opéron à l'aide de représentations moléculaires en 3D
Fonctionnement de l'opéron lactose⚓
La régulation de l'opéron lactose dépend d'une part de la présence ou non de lactose dans l'environnement, mais aussi de la présence ou non de glucose.
En effet, la synthèse des protéines impliquées dans le métabolisme du lactose serait une perte d'énergie pour la bactérie s'il n'y a pas de lactose dans l'environnement. D'autre part, le glucose reste la source d'énergie préférentielle de la bactérie car directement utilisable.
Conditions de fonctionnement⚓
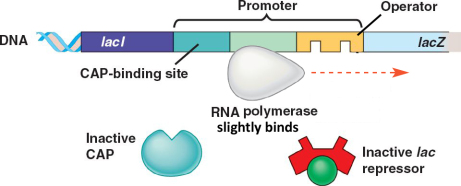
En absence de lactose⚓
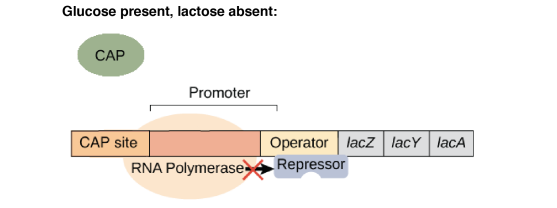
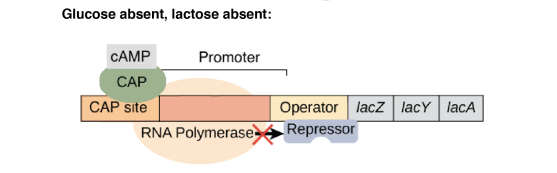
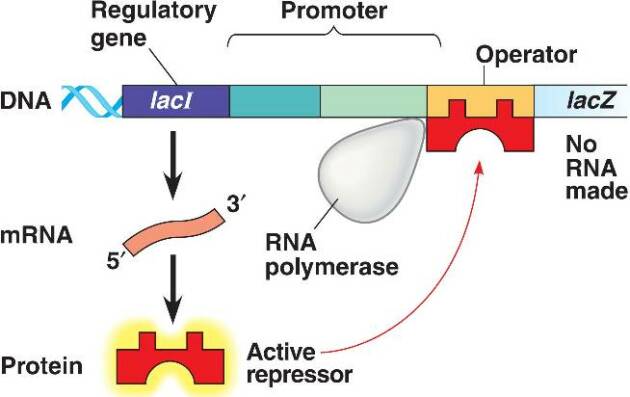
En absence de lactose, le répresseur est sous sa forme active. Il va se lier au niveau de l'opérateur de l'opéron lactose bloquant l'accès de l'ARN polymérase au site d'initiation de la transcription.
Ainsi, il y a régulation négative de la transcription des gènes de l'opéron lactose (= l'interaction empêche la transcription). Les enzymes nécessaires au métabolisme du lactose ne sont pas synthétisées car inutiles en absence de lactose.
En présence de lactose⚓
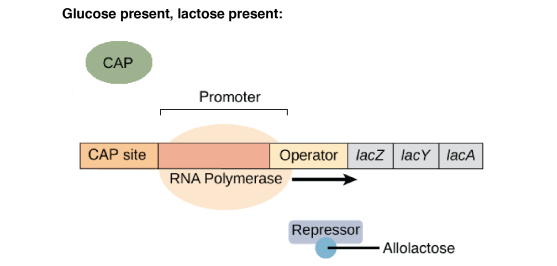
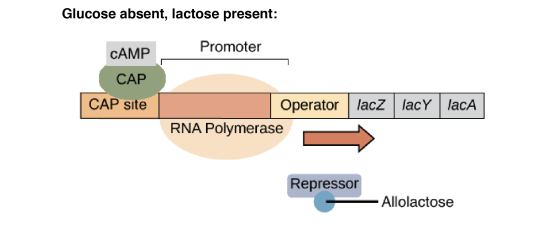
En présence de lactose, l'allolactose, un isomère du lactose, va jouer le rôle d'inducteur en se liant au répresseur. Cette liaison entraîne un changement conformationnel du répresseur qui perd alors son affinité pour l'opérateur.
Le site opérateur étant libéré, l'ARN polymérase peut atteindre le site d'initiation de la transcription et synthétiser l'ARN polycistronique. La production des enzymes nécessaires au métabolisme du lactose est donc dépendante de la présence du substrat.
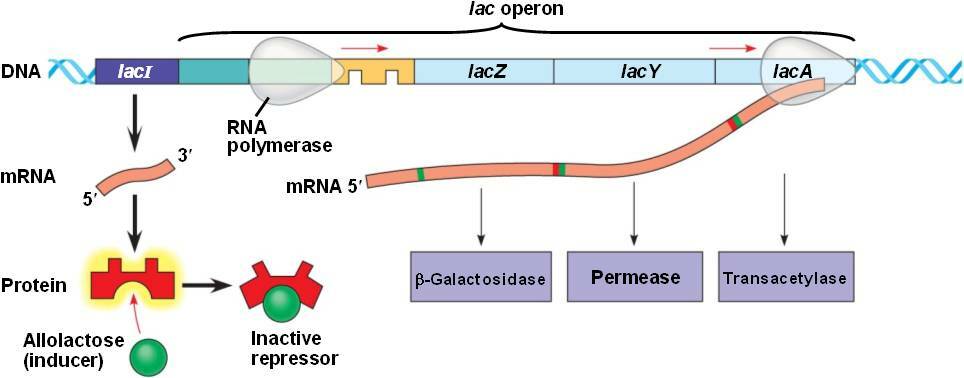
Elle a pourtant peu d'affinité pour le promoteur et la transcription est peu efficace.
Influence du glucose⚓
En présence de glucose, la bactérie va utiliser celui-ci en priorité par rapport au lactose. L'opéron lactose ne présente donc un intérêt pour la bactérie qu'en absence de glucose.
Lorsque la concentration en glucose diminue, la concentration en AMPc augmente. Celui-ci va alors se fixer à la protéine CAP (Catabolite Activator Protein, également appelée CRP pour cAMP Receptor Protein).
L'affinité de l'ARN polymérase est augmentée (50x) par la liaison de la protéine CAP-AMPc à l'ADN. On parle de répression catabolique (en présence du glucose, source d'énergie privilégiée, l'opéron lactose est peu exprimé).
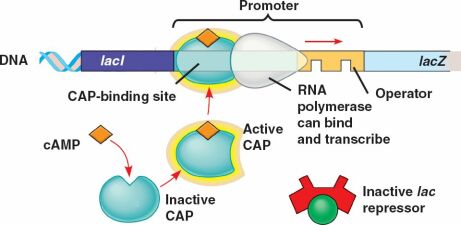
L'induction de l'opéron lactose nécessite donc deux conditions : d'une part il faut que le lactose soit présent et d'autre part que le glucose soit absent. La transcription de l'opéron lactose est donc sous le contrôle de deux protéines régulatrices :
Le répresseur lac I qui se fixe au niveau de l'opérateur en absence de lactose et bloque l'ARN polymérase. C'est une régulation négative (l'interaction inhibe la transcription).
La protéine CAP qui, sous forme d'un complexe avec l'AMPc, se lie à l'ADN et permet d'augmenter l'affinité de l'ARN polymérase pour le promoteur. C'est une régulation positive (l'interaction stimule la transcription).
Est-ce bien compris ?⚓
L'organisation en opérons présente plusieurs avantages pour la bactérie :
l'expression des différents gènes impliqués dans une même voie métabolique est coordonnée.
économie des moyens de régulation